Life History of Tristerix
aphyllus
One of the most unusual of all genera and species of
Loranthaceae
is Tristerix aphyllus. The mistletoe parasitizes
two species
of cactus, Echinopsis (=Trichocereus) chilensis
and Eulychnia acida. The mistletoe seed germinates,
forms
a radicle whose tip differentiates into an haustorium that attaches
to the host epidermis. Tissues of the haustorium then enter the
host and grown into an internal endophyte which is the highly
reduced vegetative stage of the mistletoe. The inflorescence of
the parasite is produced by adventitious buds formed on the endophyte
which then grow outward through the tissues of the cactus host.
In some cases the inflorescence axes bear small leaves, yet in
other cases these leaves are absent. Tristerix aphyllushas
been reported to be a nonphotosynthetic holoparasite (Kraus
et al. 1995), however, the presence of green tissues in seedling
radicles (see photos below) suggest that chlorophyll is present.
Moreover, the Nickrent lab has generated sequences of several
chloroplast genes indicating a chloroplast genome (plastome) is present
and functional.
Habit Photographs
- Photo.
Fray Jorge National Forest, Chile showing many Echinopsis
chilensis cactus plants infected with Tristerix
aphyllus. January 2003. Photo by G Amico.
- Photo.
Same as above - close-up of inflorescences. Photo by John W. Reynolds.
- Photo.
The mistletoe and the host (Echinopsis chilensis) in
flower. Chile. Photo by G. Glatzel.
- Photo
Mistletoe parasitic on Echinopsis chilensis, along
road heading into the Andean foothills, east of Santiago Chile. The
stem of the parasite actually grows underneath the outer cortex of the
cactus - a good way to avoid the heat! Photo by John W. Reynolds.
- Photo.
Same as above - closer view of inflorescences. Photo by John W.
Reynolds.
- Photo.
Cactus (Echinopsis chilensis) with plants. Fray
Jorge National Park, Chile. Photo by G. Amico.
- Photo.
Flowering plants on cactus. Near the Yerba Loca National Park, Chile.
Photo by G. Amico.
- Photo.
Cactus with flowering plants. Near the Yerba Loca National Park,
Chile.
Photo by G. Amico.
- Photo
Mistletoe on Echinopsis chilensis with Guillermo
Amico. In Fray Jorge National Forest, Chile. January 2003. Photo by
Mariano Rodriguez Cabal.
- Photo.
Mistletoe emerging from cactus host. Valle
Nevado, metropolitan region (ca1500 m). Photo by Serge Aubert, 9
Jan. 2003.
- Photo.
Close-up of flowers. In Fray Jorge National Forest, Chile. January
2003. Photo by G. Amico.
- Photo.
Another close-up of the flowers. Chile. Photo by G. Glatzel.
- Photo.
Echinopsis chilensis with a yellow form of Tristerix
aphyllus. Reserva Nacional Las Chinchillas, Chile. Photo
by Wilfredo Gonzáles
Lozada.
- Photo.
Closer view of the flowers of the yellow form. Reserva Nacional Las
Chinchillas, Chile. Photo by Wilfredo Gonzáles Lozada.
- Photo.
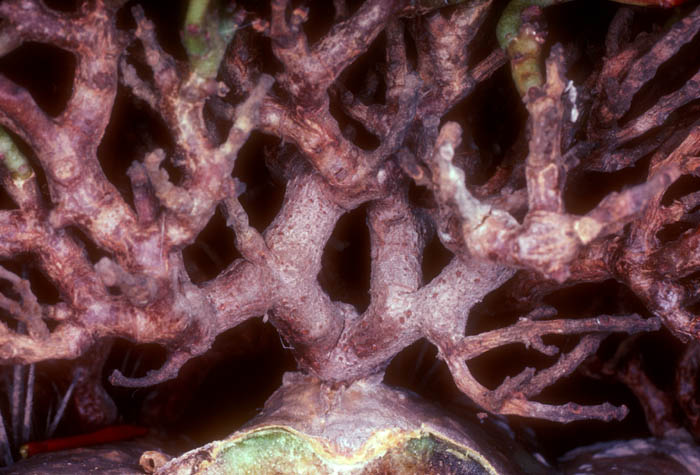
Living (top) and dead (bottom) Tristerix aphyllus
parasitic on the cactus, Echinopsis chilensis. Near
Santiago, Chile. May 1991. See Mauseth et al. (1984, 1985). Photo by
J.
Mauseth.
- Photo.
Inflorescence showing the prolific branching as it departs from its
cactus host, Echinopsis chilensis. Near Santiago,
Chile. January 1983. See Mauseth et al. (1984, 1985). Photo by J.
Mauseth.
- Photo.
Fruiting plants. Fray Jorge National Park, Chile. Photo by G. Amico.
- Photo.
Mistletoe with flowers and fruits. Chile. Photo by G. Glatzel.
- Photo.
Mature fruits containing viviparous embryos with elongated, twisted
cotyledonary petioles. Reserva Nacional Las Chinchillas, Chile. Photo
by Wilfredo
Gonzáles Lozada.
- Photo.
Another view of fruits, showing green and red embryos within. Reserva
Nacional Las Chinchillas, Chile. Photo by Wilfredo Gonzáles
Lozada.
- Photo.
Fruit with pericarp split open revealing the viscid seed and
viviparous
embryo. Reserva Nacional Las Chinchillas, Chile. Photo by Wilfredo
Gonzáles Lozada.
- Photo.
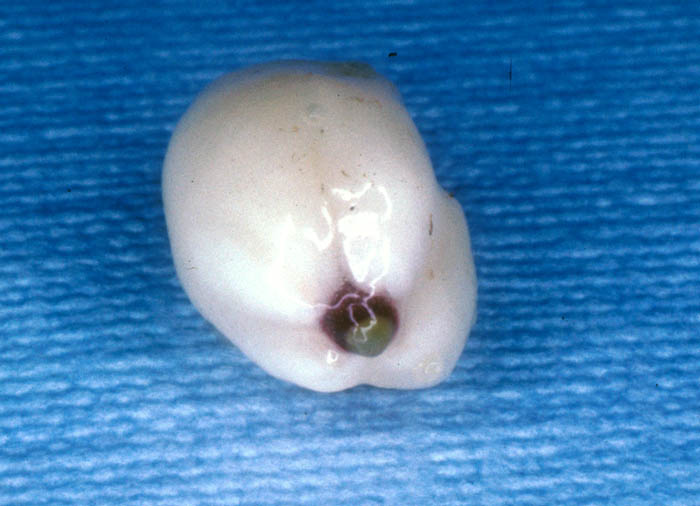
Seed with pericarp removed. The radicle with a light-colored
haustorial
disk is visible at the tip. January 1983. See Mauseth et al. (1984,
1985). Photo by J. Mauseth.
- Photo.
The seed disperser of Tristerix aphyllus, Mimus
thenca in Fray Jorge National Forest, Chile. October 2003.
Photo by Mariano Rodriguez Cabal
- Photo.
Close-up of germinating seeds on cactus. Fray Jorge National Park,
Chile. Photo by G. Amico.
- Photo.
Seedling, attached to host needle, with extremely elongated, fused
cotyledonary petioles
"reaching" for host tissue. Reserva Nacional Las Chinchillas, Chile.
Photo by Wilfredo Gonzáles Lozada. See Delrio et al. (1995)
for a discussion of the relationship between cactus spine length and
mistletoe seedling length.
- Photo.
Seedling with tape measure showing that the fused cotyledonary
petioles are ca. 10 cm long.
Reserva Nacional Las Chinchillas, Chile. Photo by Wilfredo
Gonzáles Lozada.
- Photo.
Seedlings of T. corymbosus
showing the cotyledonary petioles that are fused at their tip (inside
the seed), as in T. aphyllus and apparently other members of the
genus,
but are not fused outside the seed. In T.
aphyllus, these petioles are fused into one structure that
elongates tremendously. The epicotyl never forms in T.
aphyllus and the radicle occurs on a very short hypocotyl.
In T. corymbosus,
the epicotyl is functional such that after host attachment and primary
haustorial growth, the aerial shoots of the mistletoe develop from the
epicotyl.
- Photo.
Seedling attaching to host tissue with a primary haustorium. After
successful attachment, the cotyledonary petioles wither and
development of the
parasite endophyte occurs entirely within the host tissue. Thus, the
stems and inflorescences are not formed from the epicotyl but
endogenously from the endophyte. Reserva Nacional Las Chinchillas,
Chile. Photo by Wilfredo Gonzáles Lozada.
- Photo.
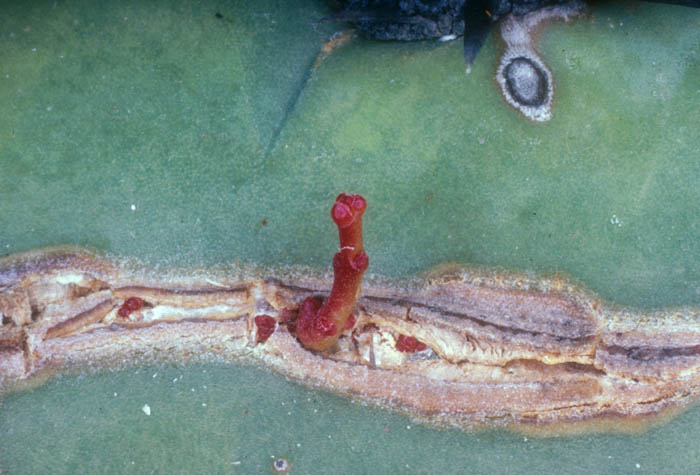
Echinopsis chilensis areole with a young floral
branch of the mistletoe emerging from its center. Near Santiago,
Chile.
October 1984. See Mauseth et al. (1984, 1985). Photo by J. Mauseth.
- Photo.
A crack in the epidermis of Echinopsis chilensis
with numerous floral buds of Tristerix emerging.
See Mauseth et al. (1984, 1985). Photo by J. Mauseth.
- Photo.
Mistletoe inflorescence emerging from Echinopsis chilensis.
Note the small leaves (bracts) subtending some of the axes.
- Photo.
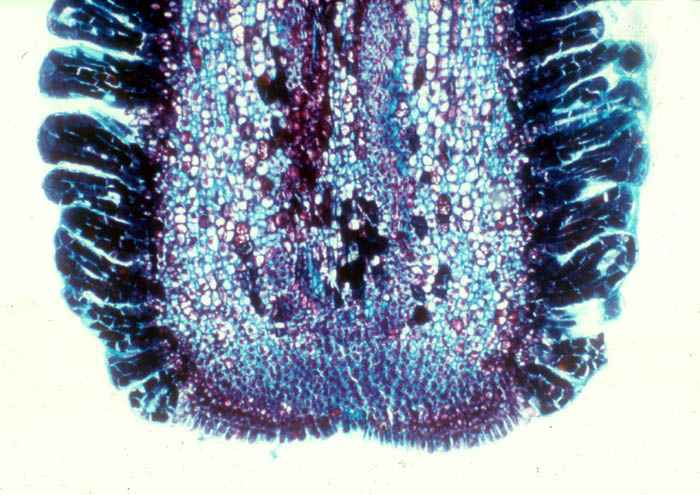
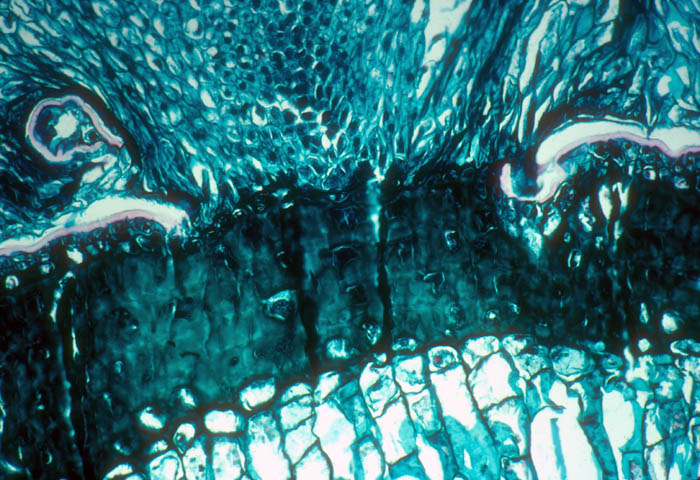
Cross-section through the cactus stem shown the haustorial strand of
the mistletoe penetrating through the cortex. Chile. Photo by G.
Glatzel.
The papers published by James Mauseth and
collaborators
(cited below) greatly increased our knowledge of the anatomy of
the infection process. I thank Jim for allowing access to his
original color slides for scanning, some of which were published
(in black and white) in the original journal articles.
Anatomy Photographs
- Photo.
Longitudinal section of elongating
radicle-haustorium of Tristerix aphyllus.
Multiseriate trichomes occur over the surface except on the haustorial
disk. See Mauseth et al. (1984, 1985). Photo by J. Mauseth.
- Photo.
Haustorium attached to Eulychnia acida
(Cactaceae). The haustorium has removed the cuticle and epidermis of
the host and two collapsed zones are visible. September 1984. See
Mauseth et al. (1984, 1985). Photo by J. Mauseth.
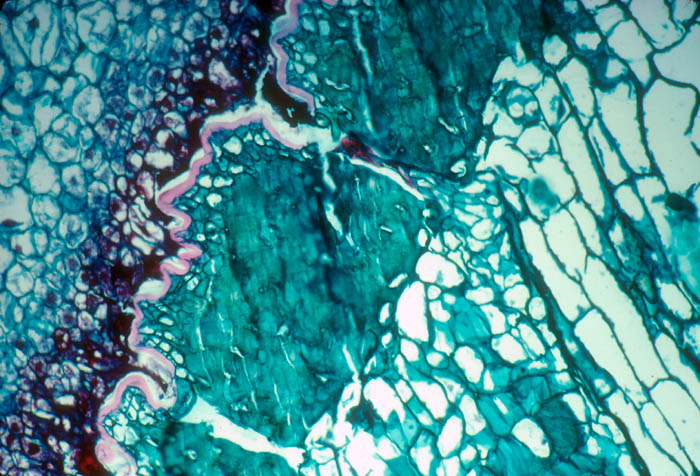
- Photo.
Transstomatal penetration of Eulychnia
acida by the haustorium of Tristerix aphyllus.
The haustorium is on the top and the host cuticle and epidermis below,
then a thick hypodermis, and at the extreme bottom host parenchymatous
cortex. The parasite filament has broken at the level of the stoma.
See
Mauseth et al. (1985) Fig. 19. Photo by J. Mauseth.
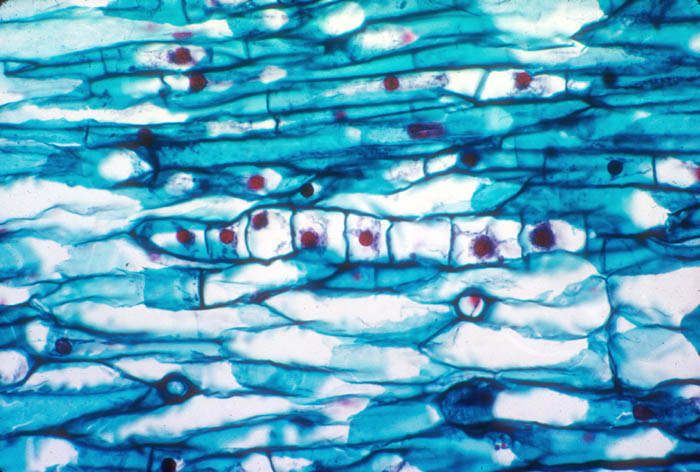
- Photo.
Hyphalike filament of the Tristerix
endophyte. The parasite has large, red-staining nuclei compared with
the host. See Mauseth et al. (1984). Photo by J. Mauseth.
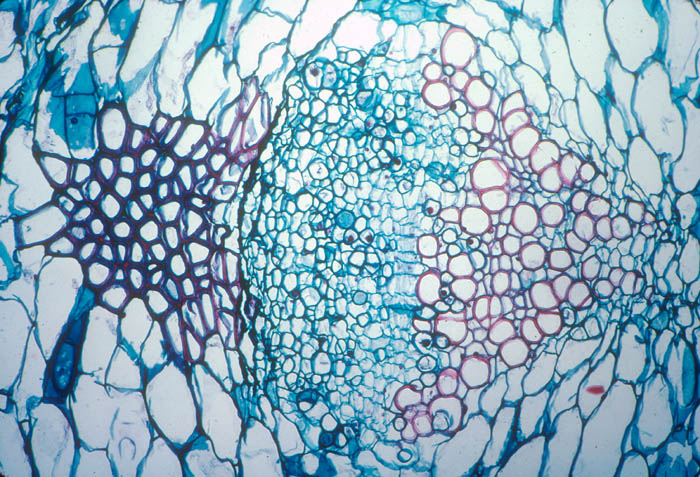
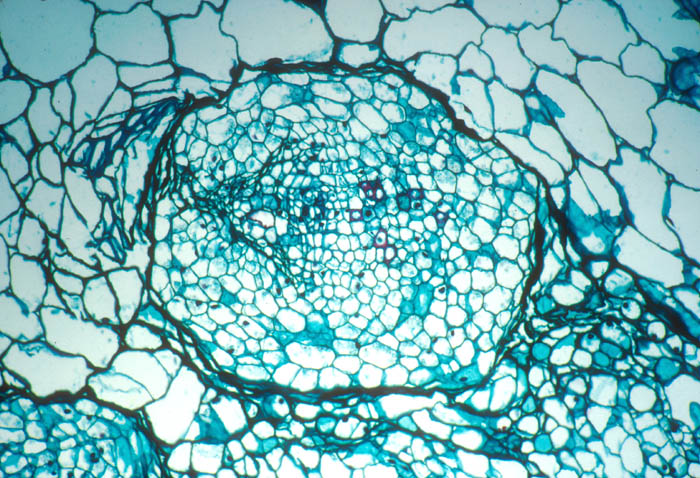
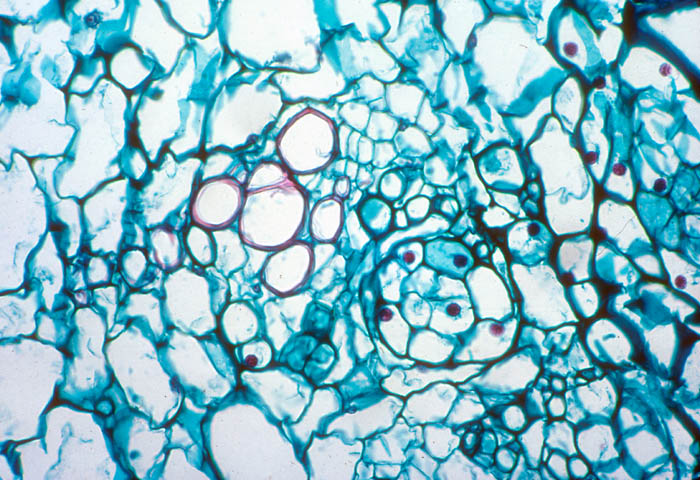
- Photo.
Vascular bundle of the cactus Echinopsis
chilensis with endophyte cells of the mistletoe (cells
scattered in central region with large, dark-staining nuclei. See
Mauseth et al. (1984). Photo by J. Mauseth.
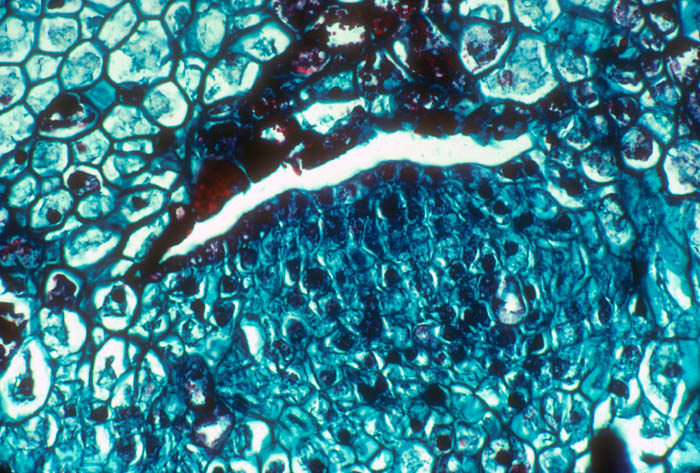
- Photo.
A mature strand of Tristerix aphyllus
growing in the phloem of a vascular bundle of the cactus Echinopsis
chilensis. A collapsed zone is just beginning to form on the
left side. The parasite cells have large, dark-staining nuclei. There
are numerous small strands and filaments of the parasite mixed in the
host phloem and cortex. See Mauseth et al. (1984). Photo by J.
Mauseth.
- Photo.
Endogenous bud forming under the epidermis of Echinopsis
chilensis. A lysigenous space has formed between the parasite
meristematic cells (below) and the host cells above. See Mauseth et
al.
(1985). Photo by J. Mauseth.
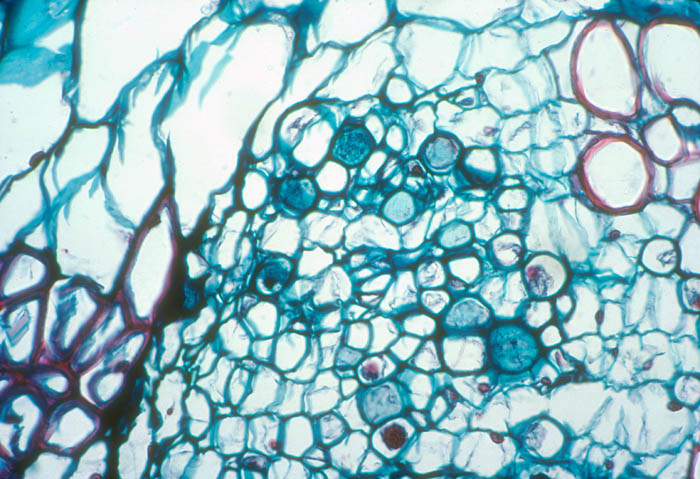
- Photo.
Magnification of the phloem of Echinopsis
chilensis infected with Tristerix aphyllus.
The parasite cells have large, dark-staining nuclei. Many of the host
seive plates are stained darker blue. See Mauseth et al. (1984). Photo
by J. Mauseth.
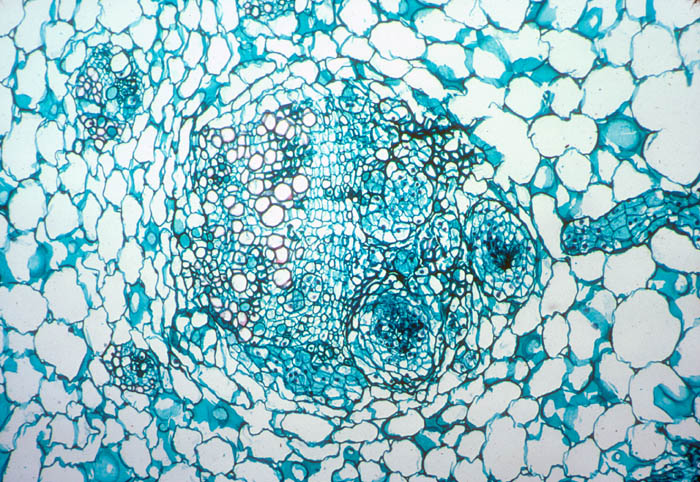
- Photo.
Echinopsis chilensis
vascular bundle in which several filaments of the mistletoe endophyte
have begun to develop into large strands. Note the strand (to the
right) that is approaching one of the axial strands. See Mauseth et
al.
(1984). Photo by J. Mauseth.
- Photo.
Early stage in the formation of the endophyte
filament of the mistletoe growing within the cortex of Trichocereus
chilensis. The parasite cells have large, dark-staining
nuclei. See Mauseth et al. (1984). Photo by J. Mauseth.
References
Amico, G. C., R. Vidal-Russell and D. L. Nickrent. 2007. Phylogenetic
relationships and ecological speciation in the mistletoe Tristerix
(Loranthaceae): the influence of pollinators, dispersers, and hosts.
American Journal of Botany 94: 558-567.
Botto-Mahan, C., R. Medel, R. Ginocchio, and G. Montenegro.
2000. Factors affecting the circular distribution of the leafless
mistletoe Tristerix aphyllus (Loranthaceae) on the
cactus
Echinopsis chilensis. Revista Chilena De Historia Natural
73: 525-531.
Caballero P, Ossa CG, Gonzáles WL, González-Browne C, Astorga G, Murúa
MM, Medel R. 2013. Testing non-additive effects of nectar-robbing ants and
hummingbird pollination on the reproductive success of a parasitic plant.
Plant Ecology 214:633-640.
DelRio, C. M., M. Hourdequin, A. Silva, and R. Medel. 1995.
The influence of cactus size and previous infection on bird deposition
of mistletoe seeds. Australian Journal of Ecology
20: 571-576.
DelRio, C. M., A. Silva, R. Medel, and M. Hourdequin. 1996.
Seed dispersers as disease vectors: Bird transmission of mistletoe
seeds to plant hosts. Ecology 77: 912-921.
Gonzáles WL, Suárez LH, Guiñez R, Mede lR. 2007. Phenotypic plasticity in
the holoparasitic mistletoe Tristerix aphyllus (Loranthaceae):
consequences of trait variation for successful establishment. Evolutionary
Ecology 21:431-444.
Gonzáles, W. L. , Suárez LH, Medel R. 2007. Outcrossing increases
infection success in the holoparasitic mistletoe Tristerix aphyllus
(Loranthaceae). Evolutionary Ecology 21:173-183.
Kelt DA, others. 2016. The avifauna of Bosque Fray Jorge National Park
and Chile's Norte Chico. Journal of Arid Environments 126:23-36.
Kraus, R., P. Trimborn, and H. Ziegler. 1995. Tristerix
aphyllus, a holoparasitic Loranthaceae. Naturwissenschaften
82: 150-151.
Lucero F, Botto-Mahan C, Medel R, Fontúrbel FE. 2014. New insights on the
mistletoe Tristerix aphyllus (Loranthaceae): interaction with diurnal and
nocturnal frugivorous species. Gayana Botánica. 71:270-272.
Mauseth, J. D., G. Montenegro, and A. M. Walckowiak. 1984.
Studies on the holoparasite Tristerix aphyllus
(Loranthaceae)
infectng Trichocereus chilensis (Cactaceae). Can.
J.
Bot. 62: 847-857.
Mauseth, J. D., G. Montenegro, and A. M. Walckowiak. 1985.
Host infection and flower formation by the parasite Tristerix
aphyllus (Loranthaceae). Can. J. Bot. 63:
567-581.
Mauseth, J. D. 1990. Morphogenesis in a highly reduced plant:
the endophyte of Tristerix aphyllus (Loranthaceae).
Bot.
Gaz. 151: 348-353.
Medel RG, Vergara E, Silva A, Serey IA. 1995. Variation of the
architectural phenotype of Tristerix aphyllus in central Chile.
Revista Chilena De Historia Natural 68:451-458.
Medel, R. 2000. Assessment of parasite-mediated selection in
a host-parasite system in plants. Ecology 81:
1554-1564.
Medel, R. 2001. Assessment of correlational selection on
tolerance
and resistance traits in a host plant-parasitic plant interaction.
Evolutionary Ecology 15: 37-52.
Medel, R., C. Botto-Mahan, C. Smith-Ramírez, M. A.
Méndez,
C. G. Ossa, L. N. Caputo, and W. L. Gonzáles. 2002. Historia
natural cuantitative de una relación
parásito-hospedero:
el sistema Tristerix-cactáceas
en Chile
semiárido.
Revista Chilena de Historia Natural 75: 127-140.
Medel, R., E. Vergara, A. Silva, and M. Kalin-Arroyo. 2004.
Effects of vector behavior and host resistance on mistletoe
aggregation.
Ecology 85: 120-126.
Midgley, G. F., J. J. Midgley, W. J. Bond, and H. P. Linder.
1994. C-3 mistletoes on CAM hosts - an ecophysiological perspective
on an unusual combination. South African Journal of Science
90: 482-485.
Silva, A., and C. M. delRio. 1996. Effects of the mistletoe
Tristerix aphyllus (Loranthaceae) on the
reproduction of
its cactus host Echinopsis chilensis. Oikos
75:
437-442.
Soto-Gamboa, M., and F. Bozinovic. 2002. Fruit-disperser
interaction
in a mistletoe-bird system: a comparison of two mechanisms of
fruits processing on seed germination. Plant Ecology
159:
171-174.
Last updated: 31-Oct-18 / dln
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}