We will begin the cycle at the time of flowering. Dwarf mistletoes, like essentially all members of Viscaceae, are dioecious, therefore staminate (male) and carpellate (female) flowers occur on different individuals. This slide of A. campylopodum (A. cyanocarpum) shows a male and a female plant parasitizing the same host tree branch. The secondary branching pattern of dwarf mistletoe shoots can be described as verticillate (whorled) or flabellate (fan-shaped), and these two types have taxonomic significance.
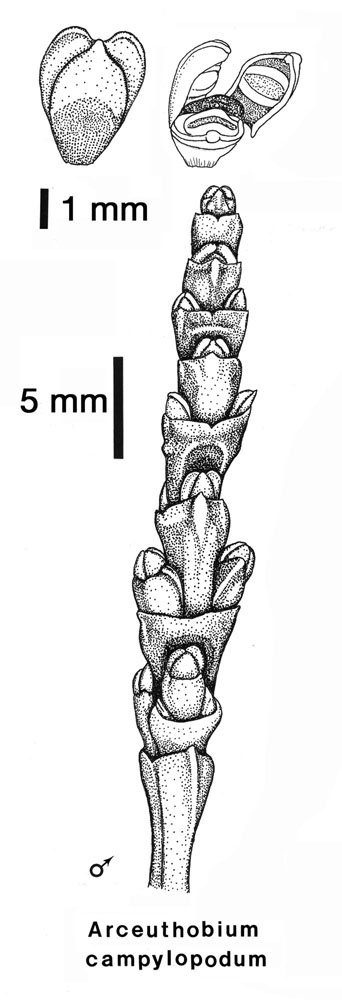
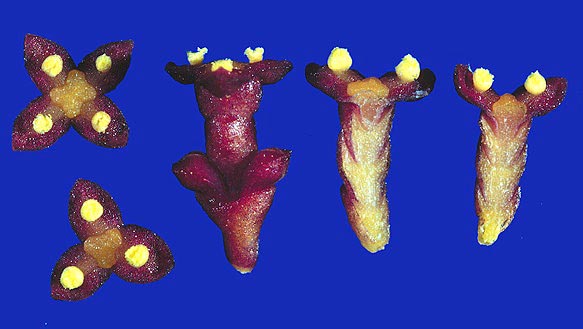
The male flowers of Arceuthobium are very reduced as demonstrated by this picture of A. pusillum or A. campylopodum. The perianth is represented only by three or four sepals, each bearing a sessile anther sac. Usually there is a prominent nectary in the center of the flower. Studies show that both wind and insect pollination occurs.
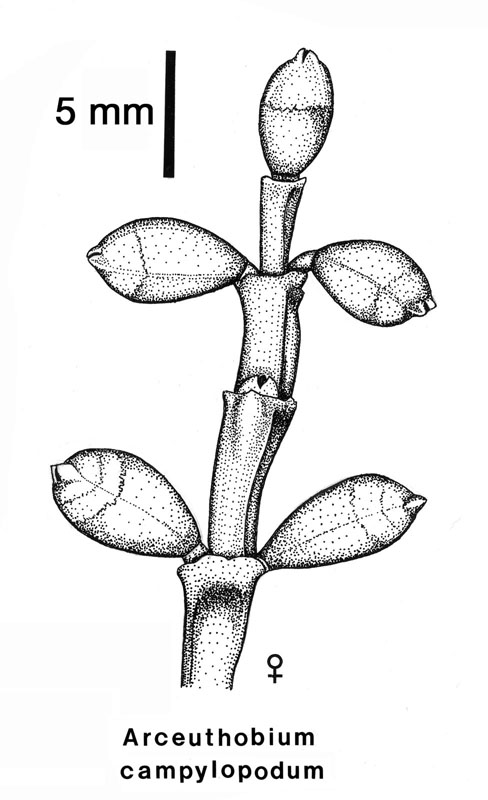
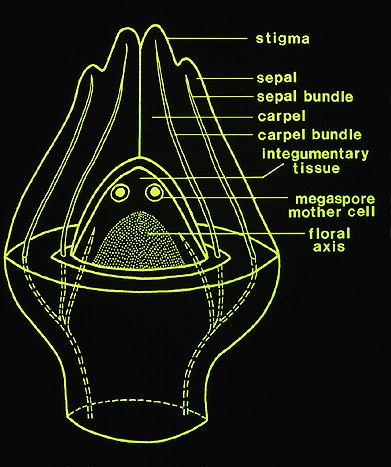
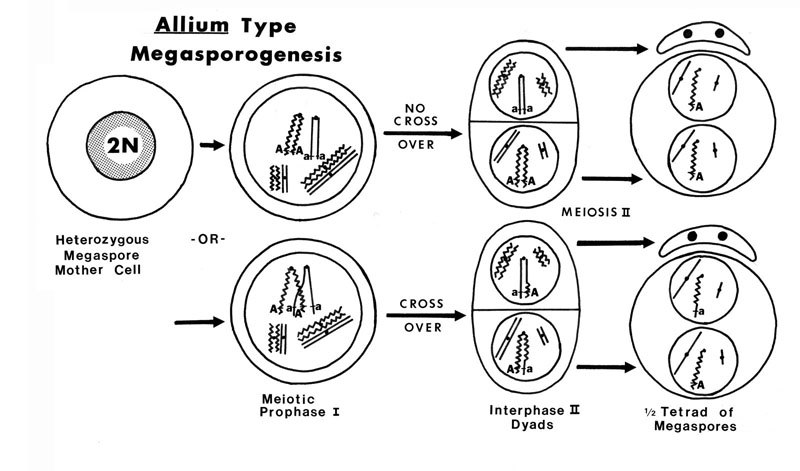
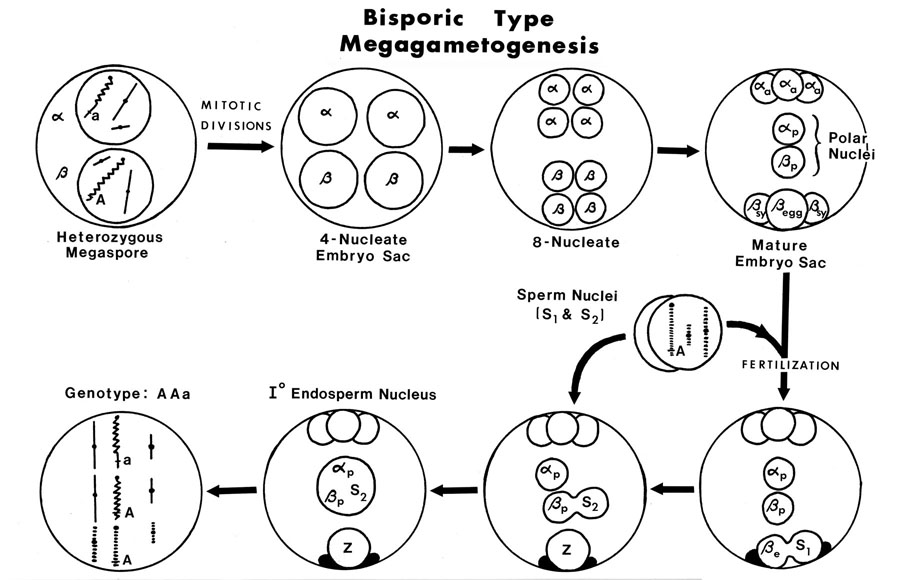
The female flowers of Arceuthobium are also very reduced as shown by this picture of A. campylopodum. Note that both flowers and fruits are present on this specimen. When mature, the female flower exudes a pollination droplet that intercepts the pollen. This is a more common syndrome in gymnosperms than in angiosperms! To fully understand the structure of the female flower, examine the diagramatic longitudinal section. The sepals are reduced and fused to the outside of the bicarpellate gynoecium. In the central part of the flower there is a space (locule) but there are no true integumented ovules. All that is present is a raised portion in the center that bears two megaspore mother cells. This mound of tissue has been called the mamelon or the placental-nucellar complex. Note that there are no integumented ovules at all. This morphology is certainly approaching the ultimate in reduction! (but check out Balanophoraceae if you want to see even smaller female flowers). Arceuthobium has an Allium type of megasporagensis. This diagram shows the two possible outcomes for a heterozygous megaspore mother cell with and without crossing over between sister chromatids. The next diagram takes the developmental process further for a heterozygous megaspore. Arceuthobium has a bisporic type of megagametogenesis, which means the mature embryo sac develops from a megaspore with two nuclei. This diagram shows the genotype of an endosperm cell which has the triploid genotype AAa.
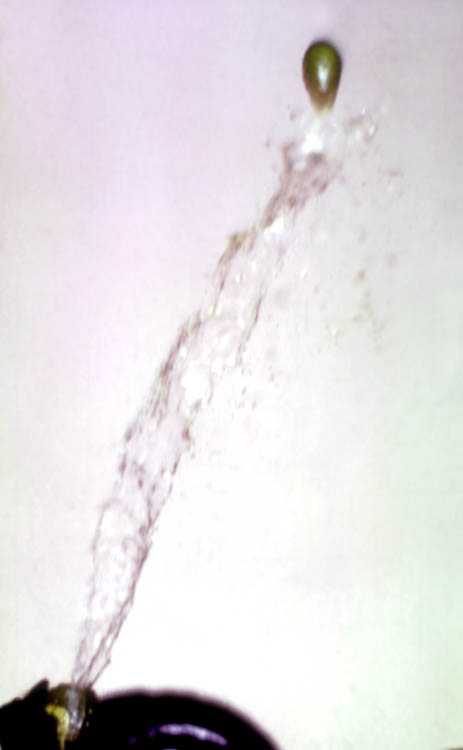
Once pollination has occurred (spring or summer, depending upon the mistletoe species), fruit development begins which may take up to a year to complete. The mature fruit is one of the engineering marvels of the plant kingdom. Unlike other mistletoes that rely upon birds for dispersal, dwarf mistletoes have evolved an explosive seed dehiscence mechanism. The fruits of all dwarf mistletoes, with the exception of A. verticilliflorum, are explosively dehiscent, one-seeded drupes (or berries) as shown by this picture of A. durangense. As the fruit matures, the pedicel begins to bend and the line that separates the pedicel from the fruit wall becomes more distinct. Hydrostatic pressure builds up within the fruit such that even the slightest touch will dislodge the fruit from the pedicel. The seed is then shot out of an opening of the rubbery fruit wall at a velocity of 27 meters per second! This movement is too rapid for the human eye to detect, but excellent high-speed strobiscopic photographs (5 millionths of a second) have been published by Hinds et al. (1963). Although most of the dispersed seeds fall within a few meters of the host crown, some travel as far as 16 meters. The number of seeds produced by one dwarf mistletoe plant is staggering. It is estimated that ponderosa pines infected with A. campylopodum may be subjected to, on average, 32,000 seeds (range 8900 to 2.2 million).
Once dispersed, the seed sticks to host needles owing to a sticky coat of viscin as shown by this slide of A. americanum on lodgepole pine. After wetting (from rain, dew), the viscin coat on the seed swells (imbibes) and becomes very slippery. This picture shows the seeds of A. vaginatum ssp. vaginatum with (right) and without (left) viscin coats. The imbibed seeds slide to the base of the needle fascicle where germination begins. Being negatively phototropic, the seedling radicle grows towards the dark which is often the underside of the branch. Only relatively young host branches can be infected. Once an obstruction is encountered, a holdfast develops on the tip of the radicle, as shown here by A. durangense or A. verticilliflorum. A penetration wedge then forms from the bottom of the holdfast which enters the host tissue. The seedling radicle can grow for some length of time since it is being nourished by the photosynthetic endosperm tissue.
Once the parasite is inside the host, the remains of the radicle and hypocotyl wither and die. This is different than other mistletoes (such as Phoradendron and most Loranthaceae) where cotyledons expand and shoot development begins from the epicotyl. The internal portion of the parasite is called the endophyte. This slide of Arceuthobium pusillum shows the cortical strands that run longitudinally along the host cambial layer and the sinkers that enter the xylem radially. The parasite tissue can be easily distinguished from the that of the host by the dark staining cytoplasm.
After a variable amount of time (months to years), a shoot apical meristem differentiates from the internal endophyte. The shoots push out through the host cortex as they grow, often marked by a swelling in the host stem. This picture shows Arceuthobium vaginatum ssp. cryptopodum on Pinus ponderosa var. scopulorum 17 months after being artificially inoculated.
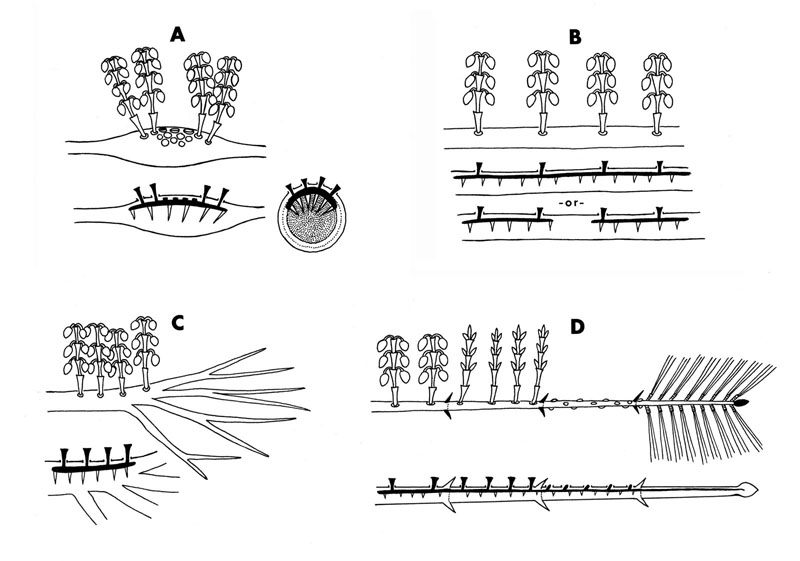
Different dwarf mistletoe species cause different kinds of infections in their hosts. A common type that is also seen in many other types of mistletoes is called a local infection. This picture of Arceuthobium campylopodum (A. tsugense) on shore pine is typical where the mistletoe endophyte is confined to a specific region of the host stem (marked by the swelling). Compare the size of the host stem distal to and proximal to the mistletoe! Two other types of infection can occur: nonsystemic and systemic brooms. Brooms (or witches brooms) result from a change in the hormone balance of the host shoots induced by a parasitic infection. Both fungi and dwarf mistletoes can induce witches brooms that are characterized by the presence of abnormal numbers of secondary branches. A nonsystemic broom occurs when a mistletoe infects a host branch, induces a broom distal to the point of attachment, but does not extensively ramify within the host tissue. Good examples of nonsystemic (anisophasic) brooms are shown in this slide of A. campylopodum (A. tsugense) infecting three different genera of host trees at one site. A systemic broom occurs when the parasite endophyte ramifies extensively within the host tissue, in some cases reaching the host shoot apical meristem. The term isophasic development is applied to those dwarf mistletoe species whose endophyte meristems are contiguous with and divide in synchrony with the host meristematic cells. Isophasic witches' brooms, as seen in A. pusillum and A. douglasii, are considered an advanced evolutionary feature. This diagram shows localized infections (A & B), a non-systemic broom (C) and a systemic broom (D).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}